
Пробелы восполнят корнями с волосатиками
Быть может, общая загадка эволюции будет раскрыта после обращения буквально к корням и истокам. Либо - через исследование жизнедеятельности настоящих паразитов. Потому как данные сферы, видимо, потенциально уже богаты сугубо научными новациями.
Во всяком случае, именно через их изучение деятели науки, как разъясняют ныне в своих пересказах специализирующиеся на открытиях науки издания, совершили пару научных открытий, которые, безусловно, помогут ученым восполнить имевшиеся до этого пробелы по эволюции.
Потому как исследователи в сопоставлении в уже многократно прокрученном учеными моменте выхода около полумиллиарда лет назад из первозданного мирового океана растений на сушу с современными растениями обнаружили новации по так называемому гравитропизму у растений, а также «точные обратные повторы в белок-кодирующих генах» в необычайно длинных митохондриальных геномах четырех видов волосатиков.
Специалисты Австрийского научно-технологического института и Педагогического университета Шэньси, как повествует в своем пересказе опубликованных в Nature Communications результатов работы исследователей «Наука и жизнь», через эксперимент сравнили, как чувствуют силу тяжести мхи, плауны, папоротники, голосеменные и цветковые растения, заставив их корневые системы расти горизонтально, наблюдая, когда корешки начнут загибаться книзу.
«Оказалось, что хуже всего из «положения лёжа» вниз растут корни у мхов, плаунов и папоротников – то есть у трёх самых древних групп наземных растений (среди которых мхи древнее и плаунов, и папоротников). Напротив, у эволюционно более молодых голосеменных (к которым относятся, например, хвойные), и цветковых (к которым относятся и пшеница в поле, и фикус на окне, и тополь на улице, и т. д.) корни довольно быстро поворачивали к низу. Иными словами, гравитропизм уверенно заработал только у голосеменных и цветковых,- разъясняется в пересказе.

Так у этих растений сработали имеющиеся в их клетках «для чувства гравитации» «особые крахмальные гранулы амилопласты, которые оседают под действием силы тяжести». Детальное рассмотрение кончика корня у разных групп растений отчетливо показало, что «только у голосеменных и цветковых амилопласты оседают прямо вниз», и «собраны» лишь «в клетках самого кончика». Тогда как «у мхов, плаунов и папоротников эти гранулы распределены по клеткам корня более равномерно», и из-за этого мхи, плавуны и папоротники - в сравнении с голосеменными - не так отчетливо, как они, чувствуют «гравитационную силу».
«С другой стороны, у голосеменных и цветковых удачно настроена передача гравитационного сигнала с помощью
гормона ауксина, который управляет ростом растительных тканей. Поток ауксина через ткани делает клетки полярными – у них появляется верх и низ. Гормон переходит от клетки к клетке с помощью белков-переносчиков, и только у голосеменных и цветковых белки-переносчики в корнях расположены в клетке так, чтобы передавать сигнал по вертикали. Рост корня регулирует не только ауксин, но, так или иначе, другие молекулярные сигналы, связанные с ауксином, тоже начинают циркулировать по клеткам корня так, что корень растёт именно вниз,- говорится в пересказе о новых результатах по гравитрапизму у растений.
При этом отмечается, что гравитропизм у наземных растений после выхода на сушу возник довольно быстро, однако «на то, чтобы отточить чувство силы тяжести, понадобились миллионы лет эволюции».
Ну, а вот российские биологи по прочтению митохондриальных геномов четырех видов так называемых волосатиков выявили в них «необычайно длинные и при этом точные обратные повторы в белок-кодирующих генах».
(Волосатики – это отдельный тип паразитоидных беспозвоночных животных, личинки коих «паразитируют на других беспозвоночных (например, насекомых и ракообразных), в то время как взрослая форма представляет собой свободно плавающее животное, напоминающее конский волос», при этом отличительная «особенность образа жизни волосатиков заключается в том, что на поздней стадии развития их личинки воздействуют на поведение своего хозяина, делая его непредсказуемым», что «рано или поздно приводит к попаданию зараженной особи в водоем», а именно это-то и требуется волосатику).
Это открытие поразило ученых, поскольку «наличие повторов в нуклеотидной последовательности накладывает сильное ограничение на возможность оптимизации закодированной ими аминокислотной последовательности в процессе эволюции», однако при этом «гены, содержащие шпильки», «тем не менее», «транскрибируются и транслируются у волосатиков в функциональные белки», поясняют в своем пересказе опубликованных в Nucleic Acids Research результатов работы российских биологов «Элементы».
До этого «митохондриальные геномы представителей одного крупного таксона — волосатиков (Nematomorpha) - оставались непрочитанными», и российские ученые вовсе не напрасно провели его читку, обнаружив удивительную и уникальную особенность.
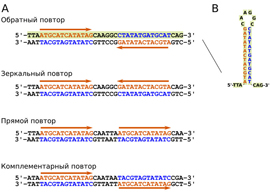
«У всех четырех видов были обнаружены длинные (до 142 нуклеотидов в одном «плече») и при этом точные обратные повторы (inverted repeat) нуклеотидных последовательностей, которые частично совпадали с последовательностями белок-кодирующих генов! Не стоит
обратные повторы (inverted repeat) нуклеотидных последовательностей, которые частично совпадали с последовательностями белок-кодирующих генов! Не стоит путать обратные повторы с прямыми, комплементарными, а также зеркальными повторами. Наличие расположенных поблизости друг от друга обратных повторов последовательности ДНК предполагает, что цепи такой молекулы ДНК могут образовывать шпильки. Шпильки формируются за счет спаривания комплементарных участков одной и той же цепи ДНК, которые образуют «стебель» шпильки. Нуклеотиды, расположенные между повторами, образуют «петлю». Формирование шпильки стабилизирует одноцепочечную ДНК и существенно меняет ее свойства
путать обратные повторы с прямыми, комплементарными, а также зеркальными повторами. Наличие расположенных поблизости друг от друга обратных повторов последовательности ДНК предполагает, что цепи такой молекулы ДНК могут образовывать шпильки. Шпильки формируются за счет спаривания комплементарных участков одной и той же цепи ДНК, которые образуют «стебель» шпильки. Нуклеотиды, расположенные между повторами, образуют «петлю». Формирование шпильки стабилизирует одноцепочечную ДНК и существенно меняет ее свойства
Повторы в нуклеотидных последовательностях ДНК широко распространены в природе и обнаружены в том числе и в других митохондриальных геномах. Так, например, точки начала репликации — участки нуклеотидной последовательности, с которых начинается репликация (копирование) молекул мтДНК, часто включают в себя обратные повторы. У бактерий обратные повторы иногда играют роль в регуляции транскрипции. Однако в подавляющем большинстве случаев повторы в нуклеотидных последовательностях не перекрываются с теми участками ДНК, которые кодируют аминокислотную последовательность белка. При наличии точного повтора одна из двух задействованных в нем последовательностей нуклеотидов полностью задает вторую. Если обе эти последовательности кодируют аминокислотную последовательность белка, то возникает плохо разрешимый конфликт: повтор накладывает значительные ограничения на аминокислотную последовательность, закодированную этими последовательностями нуклеотидов. Получается, что либо одно (точность соответствия нуклеотидов в повторе), либо другое (оптимальную для функции белка последовательность аминокислот) приходится хотя бы отчасти принести в жертву.
Поэтому нуклеотидные повторы крайне редки в тех последовательностях, которые кодируют аминокислотные последовательности — белки. Наиболее выразительным исключением из такого правила (а биология — это наука, изобилующая исключениями) являются геномы риккетсий (группа альфа-протеобактерий). В геномах риккетсий были обнаружены последовательности мобильных элементов, представляющие собой обратные повторы ДНК. Эти мобильные элементы включен в последовательность гена глутамил-тРНК синтетазы (glutaminyl-tRNA synthase) и при этом транскрибируются и транслируются — то есть в результате происходит формирование функционального белка. Однако обратные повторы в генах риккетсий не точные: на протяжении повтора некоторые отдельные нуклеотиды не комплементарны друг другу. По всей видимости, после встраивания мобильного элемента в последовательность белок-кодирующего гена мутации в процессе эволюции постепенно нарушают точность этого повтора.
В случае же с митохондриальными геномами волосатиков подавляющее большинство повторов было абсолютно точными. Во многих случаях оба «плеча» повтора кодировали консервативные (а следовательно, важные для функционирования белка) аминокислотные остатки.
Такие обратные повторы были обнаружены в большей части белок-кодирующих генов митохондриальных геномов всех четырех проанализированных видов волосатиков. Например, у волосатика Gordionus alpestris обратные повторы содержатся во всех митохондриальных генах, кодирующих субъединицы первого комплекса дыхательной цепи (комплекс I), и почти полностью перекрывают последовательность гена nad6, который кодирует одну из субъединиц. При этом оказалось, что положение некоторых обратных повторов в геномах всех четырех проанализированных видов волосатиков совпадает. Это говорит о том, что, несмотря на изменения самих нуклеотидных последовательностей, комплементарность первой и второй цепей повтора сохраняется в процессе эволюции. Однако признаков каких-либо проблем в митохондриальной энергетике у волосатиков обнаружено не было. Так, например, разрушенные клетки волосатиков были способны окислять НАДН молекулярным кислородом, и этот процесс был чувствителен к специфичному ингибитору комплекса I — ротенону. Это говорит о том, что, несмотря на необычные свойства митохондриального генома, у волосатиков работают закодированные в мтДНК ферменты,- разъясняют «Элементы».дополнительные материалыМатериализует пыль и выбросы сверхновых


При этом в пересказе подчеркивается, что в результате исследований ученым «удалось обнаружить важную особенность обратных повторов в митохондриальных геномах волосатиков».
«Дело в том, что обратные повторы, находящиеся внутри белок-кодирующего гена, могут быть трех типов, в зависимости от того, в какой «фазе» друг относительно друга находится одна и другая комплементарная последовательности. Как вы, наверное, знаете из школьного курса биологии, каждая аминокислота в белке кодируется тремя нуклеотидами в соответствии с генетическим кодом. При этом значимыми обычно являются первые два нуклеотида, а третий нуклеотид часто не несет информации. Поэтому замена третьего нуклеотида в кодирующем тринуклеотиде (кодоне) зачастую не приводит к изменению аминокислоты в составе аминокислотной последовательности. А вот изменение нуклеотида в первом и втором положении почти всегда приводит к изменению аминокислот.
В комплементарных последовательностях возможны три варианта взаимного расположения кодонов: первому положению кодона одной цепи может соответствовать первое, второе или третье положение кодона на комплементарной цепи. Эти варианты принято называть фазой 1-1, 2-2 или 3-3, исходя из положения соответствующих друг другу кодонов. С точки зрения эволюции эти фазы неравнозначны: положение 3 в кодоне редко приводит к изменениям в аминокислотной последовательности. Замена нуклеотида в положениях 2 и 1, наоборот, почти всегда меняет аминокислоту, а, стало быть, может привести к изменению эффективности работы белка.
Из этого следует, что в обратных повторах в фазах 1-1 и 2-2 сохраняется определенная степень свободы: заменяя нуклеотиды в третьем положении, можно добиться точного совпадения нуклеотидов, не изменив при этом аминокислотную последовательность, закодированную обоими комплементарными последовательностями. Таким образом, в процессе эволюции существует больше возможностей оптимизировать аминокислотную последовательность, сохраняя комплементарность обратного повтора, если эта последовательность закодирована обратным повтором в фазе 1-1 или 2-2. Обратные повторы в фазе 3-3, в которых третьи положения кодона одного «плеча» соответствуют третьему положению кодона другого «плеча», наоборот, находятся в невыгодном положении. В них мало значимые третьи кодоны находятся друг напротив друга, и та свобода, которая возможна для шпилек фаз 1-1 и 2-2, у них отсутствует.
Оказалось, что в митохондриальных геномах волосатиков длинные обратные повторы в фазе 3-3 встречаются значительно реже, чем обратные повторы в фазах 1-1 и 2-2. Этот результат косвенно указывает на то, что аминокислотные последовательности, закодированные обратными повторами в митохондриальных геномах волосатиков, находятся под давлением естественного отбора, который в большей степени благоприятствует повторам в фазах 1-1 и 2-2,- поясняется в пересказе.дополнительные материалыЭйнштейна подтвердил центр Млечного Пути
Однако деятелям науки осталось многое непонятным, у них в частности пока нет ответа на вопрос, откуда у волосатиков вообще взялась обнаруженная ими особенность.
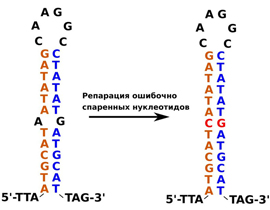
«Дело в том, что, в отличие от ядерной ДНК, митохондриальная ДНК большинства животных реплицируется асинхронно. Сначала реплицируется большая часть одной цепи ДНК, и только потом начинается репликация второй, комплементарной ей, цепи. В результате одна из цепей ДНК долгое время находится в одноцепочечном состоянии. В это время отдельные участки такой цепи могут спариваться друг с другом, образуя короткие неточные шпильки, — в достаточно длинной случайной последовательности нуклеотидов с высокой вероятностью найдутся короткие частично комплементарные элементы последовательностей.
В то же время в клетке существует система репарации ДНК, которая находит и исправляет неспаренные нуклеотиды в двуцепочечной ДНК. Стебель шпильки с точки зрения этой системы ничем не отличается от двуцепочечной ДНК, хотя и образован одной молекулой. Такая система функционирует в ядре, но есть данные о том, что подобная система репарации работает и в митохондриях. В этом случае такая система могла бы репарировать шпильки, образованные цепью митохондриальной ДНК, высвободившейся в процессе репарации, принимая их за повреждения двуцепочечной ДНК. Однако никаких экспериментальных фактов, подтверждающих такое предположение, пока что нет,- подчеркивают «Элементы».
Ученые также не обрели ясности по поводу того, какое именно «свойство волосатиков привело к тому, что у них такой механизм работает, в то время как у подавляющего большинства других видов эукариот таких повторов нет или они значительно короче».
То есть – деятелям науки еще есть с чем разобраться у волосатиков.
То есть - процесс устранения пробелов в эволюции будет, несомненно, продолжен.