
Разобрались с внутренним ухом и динозавровым полетом
Трижды могшие стать птицами динозавры оказались таковыми единожды.
Деятели науки узнают все больше подробностей о жизни динозавров, крупнейших поворотах в эволюции данных наиболее крупных в истории Земли животных.
Не так давно английские палеонтологи (из Бристольского университета), к примеру, смогли сконструировать практически точную копию мозга динозавра семейства текодонтозавров (предшественника зауроподов), довольно шустро бегавшего по планете Земля примерно 205 млн лет назад.
По копии его мозга исследователи смогли воссоздать и его внешний облик и описать его основные возможности.
Он был уже двуногим, а не четвероногим, был всеядным, употребляя и растительную пищу и охотясь на небольших животных.
Его так называемое внутреннее ухо позволяло различать довольно высокие частоты, поэтому он «обмениваться сигналами с сородичами и распознавать звуки от других животных».
Ну, а вот группа международных палеонтологов вплотную подступили к разгадке тайны про то, как динозавры подступили к активному полету, и почему, собственно, трижды пересекавшие в ходе своей эволюции физический порог способности к полету хищные динозавры только в одной своей эволюционной ветви дали настоящих птиц.
Интересно, что «претендентов на активный полет, физически способных достичь его при небольшой «доработке», было очень много — гораздо больше, чем тех, кто сумел по-настоящему полететь».
При этом ученые отмечают, что «летающие и нелетающие динозавры длительное время - никак не меньше 80 миллионов лет - жили вместе, в составе единой фауны», так как «выход той или иной группы на новый эволюционный уровень не отменяет успешности предыдущих уровней, и «конец этого сосуществования наступил только в конце мелового периода, когда вымерли все динозавры и их потомки, кроме единственной ветви эуорнисов».
О перипетиях динозаврового полета и его эволюции повествуют в своем пересказе «Элементы».
«Сразу же, как стало ясно, что птицы произошли от рептилий (а ясно это стало буквально в самый момент рождения эволюционной биологии), приобрела популярность идея, что их предки вели древесный образ жизни. Одним из первых эту точку зрения высказал американский палеонтолог Отниэл Марш (Othniel Charles Marsh; O. Marsh, 1883. Birds wih teeth). Он считал, что рептильные предки птиц («reptilian birds» — «рептильные птицы», как он выражался) были небольшими древесными существами, которые умели планировать, примерно как современные белки-летяги, летучие драконы и веслоногие лягушки. При перепрыгивании с ветки на ветку и с дерева на дерево даже зачаточные перья повышали планирующую способность, давая своему обладателю заметное преимущество. Таким образом, для «запуска» эволюции в сторону полета совершенный перьевой покров не требовался; никакой проблемы «нередуцируемой сложности» здесь не возникает. Достаточно было создать для начала хоть какие-то перья, а потом постепенно усложнять их.
Почти одновременно с Маршем другой американский палеонтолог, Самуэль Уиллистон (Samuel Wendell Williston), предложил альтернативную версию происхождения птиц — наземную. Вот что у него говорится на эту тему: «Нетрудно понять, как передние лапы динозавров могли измениться до состояния крыльев. В триасе, откуда у нас мало ископаемых находок, на протяжении долгого времени могло происходить постепенное удлинение пальцев и увеличение чешуй — и то и другое помогало животному в беге. Превратиться в перья таким чешуям было уже легко. Вероятно, поначалу крылья использовались для бега, потом для прыжков и спуска с высоты, и наконец, для парящего полета» (S. W. Williston, 1879. Are birds derived from dinosaurs?).
Уверенности Уиллистона в том, что превратиться в перья было для чешуй «очень легко», можно позавидовать. К тому же, как видим, Уиллистон все-таки допускал, что между использованием крыльев для бега и полетом был промежуточный этап в виде «прыжков и спуска с высоты». Да и высказался он об этом совсем кратко, мимоходом.Гораздо более обстоятельно наземную гипотезу происхождения птиц разработал в начале XX века венгерский палеонтолог Франц Нопча (Franz Nopcsa), человек очень яркой биографии, один из героев недавно вышедшей книги Антона Нелихова «Палеонтологи и их питомцы» (F. Nopcsa, 1907. Ideas on the origin of flight). Сравнив птиц с другими летающими позвоночными, Нопча обратил внимание на то, что ни птерозавры, ни летучие мыши не проявляют ни малейших признаков адаптации к бегу. Третья группа летающих позвоночных — птицы — образует с ними резкий контраст. Если птерозавры и летучие мыши наверняка произошли от четвероногих древесных животных, то о птицах этого сказать нельзя. По способу передвижения они гораздо ближе к наземным двуногим динозаврам. Вероятнее всего, предок птиц тоже был наземным и двуногим. И Нопча предположил, что птицы произошли от бегающих динозавров, которые научились прямо в ходе бега взлетать в воздух, используя для этого освободившиеся оперенные передние конечности.
Вывод Нопчи был слишком смелым, чтобы стать общепринятым. В то же время игнорировать большое сходство в скелете птиц и бегающих динозавров, особенно в задних конечностях, было невозможно. В результате самой популярной стала так называемая комбинированная теория: птицы полетели с деревьев, но еще до этого они прошли эволюционную стадию наземных бипедальных бегающих животных.
Развернутую версию этой теории предложил видный американский эволюционный морфолог Уолтер Бок (Walter Bock). В 1965 году он опубликовал актуальную до сих пор статью «Роль адаптивных механизмов в происхождении высших уровней организации», в которой на примере птиц рассматривал общий сценарий возникновения новой крупной группы живых организмов.
Бок считал, что крупные макроэволюционные изменения складываются из последовательности небольших шагов, на каждом из которых организм полностью адаптирован к своей текущей экологической нише. В этом смысле переходных эволюционных стадий не существует: любая из них самодостаточна. В истории происхождения птиц Бок выделяет шесть предполагаемых стадий: бипедальный бег по земле — лазанье по деревьям — прыжки с дерева на дерево — парашютирование — планирование — активный полет. Каждая стадия сопровождается специализацией в собственном направлении, которое вовсе не обязано совпадать с предыдущим и последующим, так что результирующая макроэволюционная траектория получается весьма ломаной — не как подъем по лестнице, а как путь муравья в лабиринте (это сравнение недавно использовал Эдвард Уилсон в своей замечательной книге об эволюции социального поведения).
Так или иначе, в первые две трети XX века проблема происхождения птиц, в общем, не вызывала бурных дискуссий. Но в 1970-х годах появились работы еще одного североамериканского палеонтолога — Джона Острома (John Harold Ostrom), которые произвели в этой области настоящую революцию.
Динозавровая революцияОстром во всех деталях изучил скелеты, с одной стороны, археоптерикса, а с другой — хищных динозавров, особенно некрупных и «длинноруких» (с нередуцированными передними лапами). Он показал, что сходство между ними гораздо сильнее, чем раньше думали. Поразительно похоже устроены череп, позвоночник, плечевой и тазовый пояса, кисть, стопа, и наконец — весь в целом скелет передней конечности. Последняя буквально идентична передней лапе некоторых хищных динозавров, но притом совершенно не похожа на переднюю лапу додиназавровых рептилий — текодонтов. «Анатомия археоптерикса — это почти целиком анатомия динозавра, а не текодонта, не крокодила и не птицы», — писал Остром.
Остром справедливо заключил, во-первых, что такое сходство не может быть случайным, и во-вторых, что основа двигательного аппарата динозавров и археоптерикса не могла сформироваться в древесных условиях. Археоптерикс, как и большинство динозавров, полностью бипедален. Их общие предки наверняка были двуногими животными, хорошо бегавшими по земле.
Вот с этой надежной позиции Остром и повел атаку на древесную теорию. Прежде всего он обращает внимание на то, что у археоптерикса нет ни одного скелетного признака, который можно было бы однозначно истолковать как адаптацию к древесному образу жизни. Признавая, что сценарий Бока не является невозможным, Остром тем не менее предложил свой — альтернативный.
Что, если допустить, что археоптерикс вовсе не был древесным, а был наземным бегающим хищником, охотившимся на мелкую добычу — в точности как многие динозавры, на которых он так похож по скелетным признакам? Поскольку у археоптерикса с перьями все отлично, отсюда автоматически следует, что перо появилось совершенно независимо от полета. Остром считал, что сначала перьевой покров служил для теплоизоляции. Только потом некоторые перья удлинились и приобрели дополнительную аэродинамическую функцию.
Это выглядит более-менее правдоподобно, но вызывает совершенно неизбежный вопрос: зачем, собственно, перьям было удлиняться и приобретать облик маховых перьев птиц, если животное было наземным? Понимая эту проблему, Остром предположил, что передняя лапа с увеличенными перьями помогала ловить крупных летающих насекомых, работая чем-то вроде большого сачка. Две лапы действовали как пара встречных сачков; при этом чем больше была их площадь, тем больше насекомых удавалось поймать. Именно это привело к образованию крыла, и только потом оно стало служить еще и для движения.
Надо сразу сказать, что именно эта часть построений Острома вызвала больше всего критики. При схлопывании первичных крыльев между ними неизбежно повышалось бы давление воздуха, стремящееся выбросить любой попавший туда мелкий предмет. Никакие известные животные добычу так не ловят. Если уж принимать бегающую теорию, гораздо разумнее предположить, что зачатки крыльев давали чисто локомоторные преимущества — например, служили стабилизаторами при быстром беге. Физически это вполне правдоподобно, и примерно так использует крылья современный африканский страус.
Противостояние теорийКонкуренция между древесной («from the trees down») и бегающей («from the ground up») теориями происхождения полета птиц продолжается. Сейчас она усложнена множеством недавних палеонтологических открытий, которые наверняка знакомы всем мало-мальски интересующимся (в современной популяризации палеонтологии трудно найти более ходовую тему, чем пернатые динозавры), но тем не менее пару слов о них тут сказать необходимо.
На протяжении 120 лет ключевым звеном в разработке всех гипотез происхождения птиц оставалось одно и то же существо — археоптерикс. Его регулярно переизучали, сравнивали с динозаврами, крокодилами и прочими «гадами», но других источников информации о происхождении птиц у палеонтологов почти не было.
В конце XX века положение резко изменилось. Прежде всего, в 1981 году была открыта совершенно новая группа мезозойских птиц, которая получила название Enantiornithes. Внешне они похожи на обычных птиц, но отличаются от них деталями строения скелета. Например, сочленение между двумя главными костями плечевого пояса — лопаткой и коракоидом — у энанциорнисов образовано выступом на коракоиде и выемкой на лопатке, а у обычных птиц — наоборот. Пяточный сустав у обычных птиц проходит внутри предплюсны (между разными рядами ее костей), а у энанциорнисов — между предплюсной и плюсной. Таких отличий найдено довольно много, и в сумме они позволяют весьма убедительно обосновать идею, что «птичий» тип устройства скелета сложился у энанциорнисов и у обычных птиц независимо. Недаром открывший энанциорнисов британский палеонтолог Сирил Александр Уокер (Cyril Alexander Walker) выделил их сразу в новый подкласс — это очень смелая заявка. Само название «энанциорнисы» буквально означает «противоптицы». Энанциорнисы хорошо летали, они были большой и разнообразной группой, но в конце мезозоя они вымерли вместе с динозаврами.
В 1996 году были открыты первые динозавры, несомненно имевшие перья. Джон Остром говорил, что, когда он увидел фотографии остатков этих динозавров, у него «буквально подогнулись колени»: действительно, для его гипотезы, что перо возникло независимо от полета, более наглядное подтверждение трудно было вообразить. Наличие перьев у хищных динозавров предполагалось и раньше, но теперь это стало установленным фактом. Сейчас известны уже десятки родов пернатых динозавров, причем они очень разные: от 6–7-метровых гигантов до существ размером с галку. Эти открытия дали очень много интересной информации, но попутно, как всегда бывает, породили новые вопросы.
Проблема бегающей теории в том, что многие доводы, использованные в ее поддержку, можно при желании развернуть в другую сторону. Например, у авимима, одного из первых детально описанных близких к птицам динозавров, который наверняка был оперенным, передние конечности (они же крылья) имеют длину всего в 60% от длины задних (С. М. Курзанов, 1987. Авимимиды и проблема происхождения птиц). Для полета такие крылья не годятся. А у многих других пернатых динозавров они еще короче. Специализация к двуногому бегу у рептилий отчетливо сопровождается смещением центра тяжести в сторону тазового пояса, в то время как для полета он должен находиться в области плечевого.Крылья наземных оперенных динозавров, скорее всего, действительно служили стабилизаторами при быстром беге. Но стабилизаторы по своей природе как раз должны находиться далеко от центра тяжести. В итоге, по словам нашего известного палеонтолога Александра Сергеевича Раутиана, «каждый шаг в сторону специализации к бегу удаляет от полета».
Еще один талантливый палеонтолог, Сергей Михайлович Курзанов, который описал авимима и был сторонником бегающей теории, тем не менее честно признавал, что эта теория гораздо труднее воспринимается с точки зрения здравого смысла. Это действительно так. Очевидно, что условия полета сверху вниз (с возвышенного насеста) или снизу вверх (с ровной земли) принципиально различны: в первом случае животное использует гравитационное поле как источник энергии, а во втором оно должно приложить значительное усилие, чтобы это поле преодолеть. Находясь в условиях земной гравитации, проще простого упасть вниз, но еще никому не удалось упасть вверх. Конечно, насестами, позволяющими запустить планирование, могли бы служить не только деревья, но и скалы или нависающие берега, но деревьев на Земле все-таки больше.
Для Острома было важно отсутствие у археоптерикса явных древесных адаптаций и отсутствие в палеонтологической летописи динозавров, приспособленных к планированию. Первое остается верным, а вот второе, видимо, уже нет. В самом начале XXI века был открыт микрораптор, небольшой динозавр, у которого не только передние, но и задние конечности несли перья, вполне позволяющие называть их крыльями. В 1915 году сторонник древесной теории орнитолог Уильям Биб (William Beebe) предсказывал, что в эволюции предков птиц должна найтись четырехкрылая планирующая стадия. Микрораптор «выглядит так, как будто он буквально сошел со страниц статьи Биба».
Что же касается археоптерикса, то убедительную реконструкцию способа его передвижения по деревьям недавно предложил Андрей Валерьевич Зиновьев, крупный морфолог, специалист по двигательной системе птиц, который внимательно изучил остатки археоптериксов, хранящиеся в музеях Германии. По Зиновьеву, археоптерикс карабкался по стволам деревьев с ногами, подведенными под тело (как при ходьбе по земле), фиксируясь когтями передних конечностей и вовсе не опираясь на хвост. Примерно так лазают по деревьям люди — например, сборщики плодов финиковых пальм. А по достаточно толстым ветвям археоптерикс мог просто бегать, как это делают, например, современные дятлы.
Поразительная особенность птиц — их двойная локомоторная система: передние конечности специализированы к полету, в то время как пояс задних конечностей независимо от этого и столь же глубоко специализирован к бегу. Такое сочетание уникально, и оно предполагает непростой эволюционный путь — либо сильно уклоняющийся от всех других, либо на редкость многоступенчатый, либо и то, и то сразу. С этим невозможно поспорить, но само по себе это может одинаково послужить доводом как в пользу бегающей теории, так и в пользу комбинированной. В любом случае пока что все аргументы за ту и за другую — косвенные. Проблема не в отсутствии переходных форм — их даже слишком много, — а в том, что даже если бы самый что ни на есть реальный генетический предок всех птиц попался палеонтологам, его было бы невозможно выделить среди множества других «рептильных птиц», демонстрирующих бурную эволюцию.Нельзя забывать и о том, что подавляющее большинство оперенных динозавров, находки которых так прославили современную палеонтологию, жило в меловом периоде. Они были современниками птиц и уже поэтому не могли быть их предками. Это — одна из причин, почему решение проблемы происхождения полета не стоило бы ставить в слишком прямую зависимость от картины филогенетических отношений между ископаемыми животными, тем более что эта картина до сих пор подернута рябью.дополнительные материалыШагнули и завропод и граждане десятитысячной давности
Филогенетическая чаща
Версия, принятая на сегодняшний день большинством палеонтологов, состоит в том, что птицы произошли от хищных динозавров группы Maniraptora. Именно в эту группу входит большинство пернатых динозавров. Проблема в том, что птицы рано разделились на несколько эволюционных стволов, взаимоотношения между которыми неясны. Скажем, на некоторых филогенетических деревьях посреди птиц встраиваются нелетающие существа вполне динозаврового облика. Крайняя точка зрения, принадлежащая ныне покойному Евгению Николаевичу Курочкину, состоит в том, что археоптерикс и энанциорнисы действительно произошли от манирапторных динозавров, а вот веерохвостые птицы потомками динозавров вообще не являются: их эволюционная линия независимо тянется от додинозавровых рептилий, то есть от текодонтов.
В любом случае представляется бесспорным, что приобретение «птичьих» признаков шло параллельно во множестве эволюционных линий динозавров (а если прав Курочкин, то и не только динозавров). Этот процесс параллельного приобретения «птичьих» признаков предложено называть орнитизацией динозавров, по аналогии с происходившей в истории Земли несколько раньше маммализацией зверообразных. Есть и другие примеры подобных «-заций»: ангиоспермизация голосеменных растений (В. А. Красилов, 1989. Происхождение и ранняя эволюция цветковых растений), артроподизация древних линяющих животных. Вполне может быть, что именно так и выглядит универсальный сценарий происхождения крупных групп организмов — по крайней мере, в пределах эукариот.
Как сказал известный исследователь динозавров Ринчен Барсболд, «птицеподобие в эволюции поздних хищных динозавров... было неравномерным, но почти всеобщим». Большинство хищных динозавров было затронуто процессом орнитизации, хотя и не все они в итоге взлетели.
Терминологическое отступление
Принимая во внимание весь этот контекст, взглянем на новую обстоятельную статью о проблеме происхождения полета, вышедшую в октябре 2020 года и подписанную группой ученых из США, Китая, Канады и Аргентины. Среди них известные палеонтологи Марк Норелл (Mark Norell) и Стив Брусатти (Stephen Louis Brusatte), книга которого недавно вышла на русском языке (она называется «Время динозавров»). Тут можно быть уверенными, что мы имеем дело с современным «мэйнстримом» в данной области науки.
Прежде всего, однако, нам необходимо договориться о терминах. В современной систематике живых организмов, в том числе и ископаемых, господствует кладистика — направление, основанное немецким биологом Вилли Хеннигом (Willi Hennig), в котором система строится только и исключительно на родстве (доказанном или предполагаемом). Анатомические сходства и различия, образ жизни, эволюционный успех и все прочие показатели никакого самостоятельного значения для такой системы не имеют. Единственный эволюционный процесс, который кладистика признает значимым — это разделение ветвей (дивергенция), причем постулируется, что делиться они могут только надвое. Иные критерии, кроме порядка ветвлений эволюционного древа, кладистическая система не учитывает. Группы, возникшие в результате одного ветвления, называются сестринскими, и им всегда придается равный систематический ранг. И, что для нас очень важно, любая такая группа должна включать всех без исключения потомков своего последнего общего предка, независимо от того, насколько сильно они изменились по сравнению с ним.
Например, человек с точки зрения кладистики — не потомок человекообразных обезьян, а просто человекообразная обезьяна, поскольку он, несомненно, входит в эту эволюционную ветвь. Точно так же птицы, коль скоро они произошли от манирапторных динозавров, считаются просто манирапторными динозаврами, а наземные позвоночные, происходящие от костных рыб — просто одной из групп костных рыб. В статьях и книгах, написанных сторонниками кладистики, часто встречаются обороты вроде «костные рыбы, в том числе человек, мышь, цыпленок и лягушка» или «самый маленький динозавр — это колибри-пчелка, которая живет только на Кубе». В кладистике принципиально невозможно, чтобы от некоторой уже существующей группы произошла новая группа того же ранга, например от отряда — новый отряд или от класса — новый класс. Как бы сильно организм ни изменился, он все равно навеки останется в пределах своей эволюционной ветви, а значит, и в пределах охватывающего эту ветвь таксона. Любая сколь угодно масштабная эволюция с точки зрения кладистики приводит только к дроблению таксонов, ранги которых с каждым ветвлением эволюционного древа понижаются.
Это теоретическое отступление понадобилось вот зачем. С кладистической точки зрения птицы — одна из групп динозавров, поэтому считается терминологически неправильным говорить о динозаврах, не включая в это понятие птиц. Для обозначения динозавров, не являющихся птицами, введен специальный термин «нептичьи динозавры» (nonavian dinosaurs). В этом случае, например, поставленная Уолтером Боком проблема происхождения новых крупных таксонов становится совершенно бессмысленной, поскольку никаким новым таксоном, сравнимым по рангу с динозаврами, птицы не являются.
В данной статье термины «динозавр» и «птица» будут исключительно для удобства использоваться в традиционном значении: первое — для обозначения явно нелетающего существа, второе — для обозначения явно летающего. Пограничные случаи, которые нам в первую очередь как раз и интересны, будут оговариваться особо.
Полет: много званых
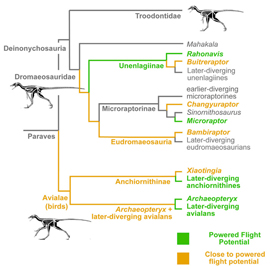
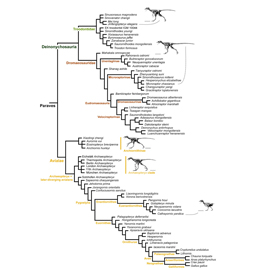
Итак, для начала посмотрим на эволюционное древо, составленное Брусатти, Нореллом и их коллегами. На этом древе присутствуют не все динозавры, не все ящеротазовые динозавры (тут надо учитывать небольшой парадокс: птицы произошли от ящеротазовых динозавров, а не от птицетазовых) и даже не все манирапторы; такое ограничение понятно, ибо нельзя объять необъятное. Перед нами древо относительно небольшой группы, которая находится внутри Maniraptora и называется Paraves. Это — птицы с их ближайшими нелетающими родственниками. В частности, именно в группе Paraves сильнее всего проявилась общая для многих хищных динозавров тенденция к постепенному уменьшению средних размеров тела, ставшая важной предпосылкой к полету. По словам палеонтолога Даррена Нэйша (Darren Naish), все нелетающие Paraves в общем похожи на короткокрылые версии археоптерикса.
С систематической точки зрения в группу Paraves входят семейства троодонтид и дромеозаврид, но не входят овирапториды, к которым относится упомянутый выше авимим и которые тоже очень похожи на птиц как минимум по ряду особенностей скелета. Дромеозавриды — очень разнообразное семейство. К ним относятся и знаменитые грозные хищники дейноних и велоцираптор, и гигантский для этой группы шестиметровый ютараптор, и совсем небольшой четырехкрылый микрораптор — он здесь тоже уже упоминался, и нам стоит обратить на него особое внимание. Троодонтиды чуть менее разнообразны, зато еще более птицеподобны.
Продолжая рассматривать древо, мы видим, что группа Paraves делится на две эволюционные ветви. Первая ветвь, состоящая из дромеозаврид и троодонтид, называется Deinonychosauria. Вторая ветвь — «птичья» — называется Avialae. В кладистической системе, о которой мы сейчас говорим, все птицы относятся к авиалам, но отсюда отнюдь не следует, что все авиалы — птицы.
Сама гипотеза о существовании ствола Maniraptora, который делится на ветви Deinonychosauria и Avialae, довольно старая: Жак Готье (Jacques Armand Gauthier) высказал ее еще в 1986 году, когда и появились все эти таксономические понятия. Норелл, Брусатти и их коллеги фактически просто подтвердили гипотезу Готье, аккуратно интегрировав в нее новые палеонтологические данные, которых, понятно, за 34 года накопилось очень много. Альтернативная версия, согласно которой троодонтиды ближе к птицам, чем дромеозавры, тоже рассматривалась, но после кладистического анализа была сочтена маловероятной. Итак, получается, что Avialae, то есть «птицы в широком смысле» — самостоятельный эволюционный ствол.
Кто, собственно, к этим «птицам в широком смысле» относится? Вновь посмотрим на древо. В первую очередь мы видим там две крупные ветви: энанциорнисы и настоящие птицы (Euornithes). Последние — единственные авиалы, дожившие до современности. Рядом с их развилкой от древа птиц отходит еще несколько небольших ветвей, в том числе необычный, но достаточно хорошо изученный конфуциусорнис, которого Курочкин считал еще одним эволюционным стволом того же ранга, что энанциорнисы и настоящие птицы. Чуть подальше находится ветвь археоптериксов, легендарных переходных форм, известных с XIX века. На самом деле нынешний несравненно более полный материал делает проблемой не поиск переходных форм — они там сплошь, — а совсем наоборот, выделение тех, которые переходными уже (или еще) не являются. Наконец, от самого основания древа авиал отходит ветвь анхиорнисов, очень интересных небольших оперенных существ, которые выглядят чуть более далекими от современных птиц, чем археоптериксы, и жили на несколько миллионов лет раньше них.
Кто же из этих животных потенциально мог бы освоить полет? Самый простой ответ: да все. Главным эволюционным «изобретением», которое дало птицам возможность полететь, является контурное перо — сложная конструкция из стержня и бородок с крючочками, смыкающихся в сплошную плоскость. У Paraves с контурными перьями не было никаких проблем. Более того, существует более крупная группа, объединяющая всех Paraves с их родственниками, тоже имевшими контурные перья. Эта группа даже название получила в честь контурного пера: Pennaraptora (penna — по-латыни перо, в том числе писчее). В группу Pennaraptora вместе с паравесами входят и овирапториды (но не входят более примитивные манирапторы, такие, например, как теризинозавры). Это означает, что перьевая «оснастка» потенциально позволяла полететь даже авимиму. Мешали другие показатели, в первую очередь пропорции тела и характеристики мускулатуры.
Полет: мало избранных
Переходя непосредственно к полету, надо прежде всего обратить внимание на две особенности обсуждаемой работы. Во-первых, на древе, построенном ее авторами, птицы строго монофилетичны: гипотезы о том, что разные группы птиц независимо произошли от нелетающих динозавров (не говоря уж о текодонтах), здесь не поддерживаются. Пару десятков лет назад даже в работах убежденных сторонников кладистики публиковались эволюционные деревья, на которых между разными эволюционными стволами птиц вклинивались заведомо нелетающие наземные динозавры, и следует признать, что концепция орнитизации динозавров со своей стороны делала такой сценарий правдоподобным. Но сейчас он непопулярен.
Во-вторых, Норелл, Брусатти и их коллеги придерживаются «бегающей» теории происхождения полета. Другие версии они не только не обсуждают, но даже и не упоминают, очевидно, считая этот вопрос давно и окончательно решенным. Тут уместно вспомнить, что в 1986 году в Калифорнии вышел замечательный сборник, озаглавленный «Происхождение птиц и эволюция полета», в котором рядом были помещены статья Уолтера Бока «Древесное происхождение полета птиц» и статья Джона Острома «Бегающее происхождение полета птиц». Сейчас в этой области плюрализма гораздо меньше.
Теперь — собственно, о новых фактах, добытых в нынешнем исследовании. Для оценки способности к полету были выделены два параметра: удельная нагрузка на крыло и подъемная сила. Их примерные величины вычислялись, естественно, исходя из морфологических данных, которых сейчас доступно много. Сочетание этих показателей, физически позволяющее перейти к активному полету, авторы называют порогом активного полета (powered flight threshold).
Когда наши авторы наложили сумму результатов расчетов на филогенетическое древо, у них получилось, что представители группы Paraves пересекали порог активного полета минимум трижды, а скорее всего — четырежды. К первой из ветвей, перешедших этот порог, относится уже знакомый нам микрораптор. Ко второй — рахонавис (Rahonavis), во многом похожий на микрораптора, но относящийся к другой подгруппе дромеозавров (впрочем, окончательно его филогенетическое положение не установлено). И к третьей — собственно птицы, с которыми, однако, в свою очередь дело обстоит не так-то просто.
Мы уже знаем, что на древе, построенном авторами обсуждаемой статьи, группа Avialae распадается на две ветви. К одной из них относятся анхиорнисы — существа, в целом похожие на археоптерикса, но чуть более древние и примитивные. Ко второй относятся все остальные птицы, включая и археоптерикса, и конфуциорнисов, и энантиорнисов, и эуорнисов (как мы знаем, до современности дожили только последние). Так вот, по данным наших авторов, анхиорнисы порог активного полета пересекли, но... не все. Как минимум один из них, а именно сяотингия (Xiaotingia), остался за порогом активного полета, хоть и близко к нему. Это должно означать, что и общий предок анхиорнисов и всех остальных птиц — то есть общий предок Avialae — порога активного полета еще не перешел. Анхиорнисы совершили этот переход независимо от остальных Avialae. Вот она — орнитизация. Даже кладистическая методология, последовательно стремящаяся к игнорированию любых эволюционных процессов, кроме дивергенций, все-таки привела к тому, что факт параллельной орнитизации пришлось признать (хотя авторы и не используют этого термина).
Следует заметить, что сяотингия до такой степени похожа на археоптерикса, что поначалу ее и сочли всего лишь китайским представителем семейства археоптериксов. С этой точки зрения важно не столько то, что в обсуждаемой статье сяотингия и археоптерикс отнесены к разным ветвям авиал — они все равно занимают и там, и там положение, очень близкое к общему корню, — сколько то, что они оказались по разные стороны порога активного полета. Археоптерикс, по данным наших авторов, пересек этот порог, а очень похожая на него сяотингия еще нет. Этот факт наглядно показывает, что орнитизация происходила чрезвычайно постепенно.
Порог активного полета — это типичная града, эволюционный уровень, который может достигаться независимо разными эволюционными ветвями (как правило, более-менее родственными). Последовательность град в происхождении млекопитающих была прекрасно показана Леонидом Петровичем Татариновым в его классических исследованиях. О филогенетическом положении однопроходных млекопитающих). Было бы просто удивительно, если бы эволюционный сценарий происхождения птиц не имел ничего общего со сценарием происхождения млекопитающих. Но, судя по всему, он выглядит как раз аналогично.
Как обычно и бывает в подобных случаях, динозавры приближались к порогу активного полета широким фронтом, экспериментируя в разных направлениях. Эта тенденция охватила не только группу Paraves. Авторы отмечают, что и у тех оперенных динозавров, которые в нее не входят (таковы, например, орнитомимиды и компсогнатиды), подъемная сила крыльев бывает не меньше, чем у современных нелетающих птиц, таких, как галапагосский баклан и попугай какапо. Кстати говоря, первые найденные пернатые динозавры — те самые, которые поразили Джона Острома — относились именно к компсогнатидам, а не к паравесам. Так что претендентов на активный полет, физически способных достичь его при небольшой «доработке», было очень много — гораздо больше, чем тех, кто сумел по-настоящему полететь. И, конечно, летающие и нелетающие динозавры длительное время — никак не меньше 80 миллионов лет — жили вместе, в составе единой фауны: выход той или иной группы на новый эволюционный уровень не отменяет успешности предыдущих уровней. Конец этого сосуществования наступил только в конце мелового периода, когда вымерли все динозавры и их потомки, кроме единственной ветви эуорнисов,- разъясняется в пересказе.дополнительные материалыНо заблуждения динозавров нейросети не переняли

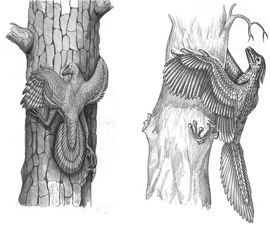


Таков пока взгляд деятелей науки на то, как в ходе своей эволюции динозавры подступались к способности летать.
Эту эволюционную картину, без сомнения, дополнят и новые открытия.